
1811-5209/23/0019-0289$2.50 DOI: 10.2138/gselements.19.5.289
Keywords: Large igneous province; magmatism; terrestrial; vegetation; palynology; mass extinction.
INTRODUCTION
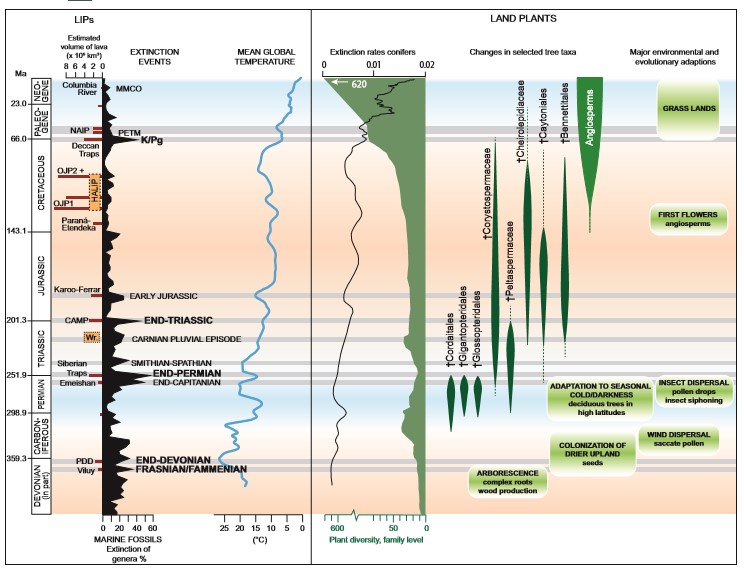
Historical and present-day volcanic eruptions, such as the 1783–1784 Laki fissure eruptions, the 1815 Tambora eruption, and the 1991 Mount Pinatubo eruption, are testaments to how volcanic activity can cause environmental and climatic changes on local to global scales. In these cases, it was primarily emissions of sulfur dioxide and volcanic ash or dust injected into the stratosphere, which blocked incoming solar radiation, that caused climatic deterioration in the year(s) following the eruptions, resulting in global cooling (Mather 2015). In contrast, the magmatic activity of large igneous provinces (LIPs) is known to have emitted vast amounts of greenhouse gases (e.g., CO2 and methane) as well as sulfur dioxide, halocarbons, and heavy metals, which can be traced in the sedimentary record and used as proxies for the volcanic activity (e.g., Lindström et al. 2021). These volcanic emissions set off a cascade of events that led to prolonged intervals (hundreds of thousands of years) of global warming, ocean nutrification, anoxia, sea-level change, ocean acidification, global cooling, wildfires, and introduction of toxic metals and gases to the atmosphere numerous times throughout the Phanerozoic. The formation of LIPs is contemporaneous with most mass extinctions and smaller-scale biotic crises throughout geological time, and LIP activity is implicated to have contributed to at least three (e.g., end-Permian, end-Triassic, Cretaceous–Paleogene), if not all, of the “Big Five” mass extinctions (Fig. 1). Study of the environmental effects of LIPs improves the understanding of the complex interactions between the geosphere, atmosphere, hydrosphere, and biosphere, and therefore the accuracy of predictions of ecosystem response to future change associated with unabated greenhouse gas emissions and metal pollution.
Terrestrial vegetation and ecosystem dynamics are an integral part of the global carbon cycle and the composition and extent of terrestrial vegetation play an important role in global climate feedback mechanisms. However, compared with the marine fossil record, the influence of LIPs on terrestrial ecosystems is not as well understood (Bond and Sun 2021). Extinctions in the animal record during major biotic crises in Earth history are not mirrored by comparable major changes in land plants (Fig. 1). For example, across the largest animal extinction event in Earth’s history, the end-Permian event (~251 Ma), few plants became extinct globally, even though this was arguably the most severe event for land plants (Nowak et al. 2019). Major changes in vegetation took place on a regional to supra-regional scale and the terrestrial crisis was underway several tens to hundreds of thousands of years before the marine extinctions (e.g., Hochuli et al. 2016; Dal Corso et al. 2022 and references therein). For the end-Triassic event, vegetation changes may have started earlier than the marine extinctions, but the major changes in both realms appear to have largely coincided (Lindström et al. 2021; Lindström 2021). During both the end-Permian and end-Triassic events, ecosystem changes and extinctions were protracted over 10s to 100s of thousands of years. In contrast, the terrestrial and marine extinctions during the Cretaceous–Paleogene event, with the effects of the Chicxulub impact, were simultaneous and of comparatively short duration (e.g., Vajda and Bercovici 2014).
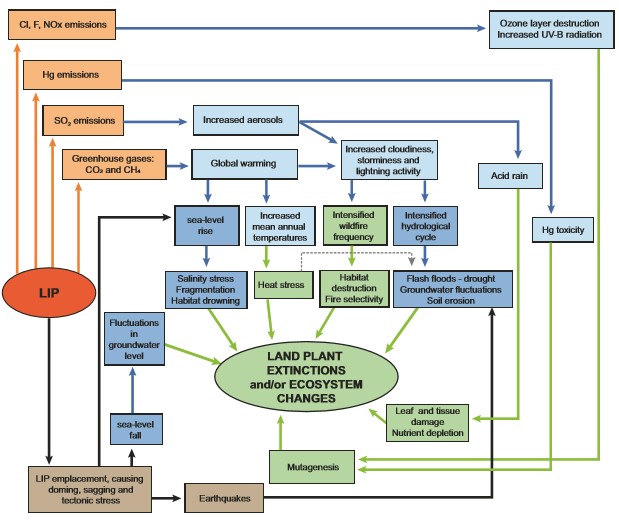
Land plants, despite being sedentary organisms, may, as a group, be more resistant to mass extinction than animals. Since their dawn, land plants have evolved adaptations to adverse changes in their environment, which provide autoecological (i.e., the way plants interact with the total environment) advantages compared with animals. These include, for example, highly resistant and dispersable resting stages (e.g., resistant seeds), various regenerative adaptations such as subterraneous structures, multiple reproductive strategies, morphological and reproductive adaption to disturbance, complex genomes, and even DNA repair mechanisms. Despite the remarkable resilience of land plants to adverse changes in the environment, LIPs still affected land plant communities in a myriad of ways: from the destruction of vegetation by lava flows in their immediate surroundings to more widely-distributed stressors, such as climate warming from CO2 emissions, short-term cooling due to SO2 emissions, sea-level change, acid rain, wildfires, photosynthesis inhibition from the release of aerosols, increased UV-B radiation due to thinning of the ozone layer by halocarbons released from magma intruding and heating evaporites, and poisoning due to release of toxic pollution including polycyclic aromatic hydrocarbons (PAHs), mercury (Hg), and heavy metals (Fig. 2). Alone or cumulatively, these and other effects of LIPs have caused destabilization of terrestrial plant communities, as recorded by fossilized pollen and spores in the rock record. Understanding the response of land plants to large-scale magmatic events offers analogues for the global consequences of modern deforestation and diversity loss (Vajda et al. 2020).
Palynology (Greek, palúnō, to strew or sprinkle) includes the study of dispersed pollen and spores from land plants. The palynological record can offer a more comprehensive view of land plant change over time than other techniques used to reconstruct past vegetation because the morphological characteristics of pollen and spores permit their identification to a low taxonomic level (often species), they are often exceptionally well-preserved, and they occur in great abundance in sedimentary rocks, permitting statistical analyses and representation of rare types. For example, the pollen and spore record often indicates greater severity in both taxonomic extinctions and ecosystem structure effects during biotic crises in the geological past than macrofossil records (Lindström 2016; Nowak et al. 2019), indicating that many of the plants that disappeared during biotic crises, or shortly thereafter, were rare plants or plants with little macrofossil preservation potential (Lindström 2021). Further biasing the record may be that the majority of the terrestrial fossil records are from lowland depositional settings, such as lacustrine or riparian environments where sediments accumulate. Thus, the macrofossil record of terrestrial land plants may be biased toward those that grew in wetlands. In contrast, the palynological record includes data from widely dispersed pollen and spores, including those from drier and/or upland habitats mixed with those from slope or lowland settings, and thus shows a more muted extinction intensity, for example, across the end-Permian extinction event (Dal Corso et al. 2022). In this contribution, we discuss the impacts of LIP magmatism on terrestrial land plant ecosystems by focusing on palynological evidence for land plant change during major biotic crises associated with LIP activity.
HOW LIP VOLCANISM IS MANIFESTED IN THE SPORE AND POLLEN RECORD
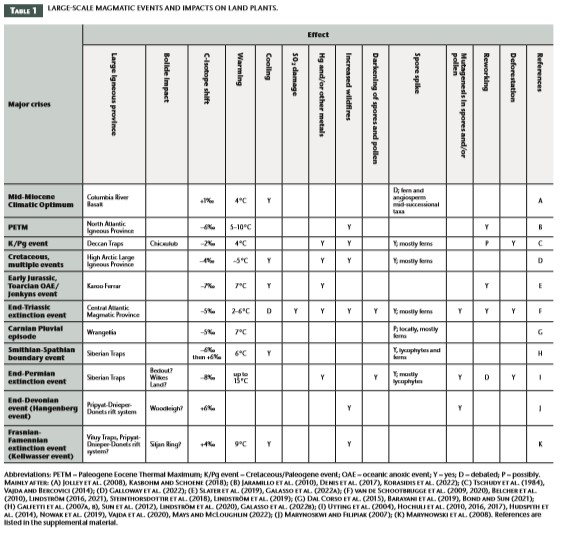
Although major changes in the evolution of land plants generally do not coincide with the Big Five mass extinctions (Fig. 1), there is ample evidence for regional-scale land plant extinction and community reorganization during almost all major crises, particularly those associated with LIP magmatism (Fig. 2; Table 1). Land plant compositional changes as recorded by pollen and spores occurred associated with the emplacement of the Siberian Traps, the Central Atlantic Magmatic Province (CAMP) the Karoo Ferrar LIP, the Kerguelen Plateau, Ontong Java Plateau, ParanáEtendeka Province, the High Arctic Large Igneous Province (HALIP), the North Atlantic Igneous Province (NAIP), and the Columbia River Basalts (Fig. 1; Table 1). While floral turnover is documented in the terrestrial fossil record at both high and mid-latitudes during most major biotic crises in Earth’s history, and in a variety of settings, the patterns of change appear to have been diachronous across latitudes (Vajda and Bercovici, 2014; Lindström 2016). More direct effects of large-scale volcanism will also differ geographically. Direct effects, such as landscape disturbance and lava-ignited wildfires, will be greater proximal to the location of lava emplacement. Conversely, atmospheric changes due to greenhouse gases and volcanic dust increase cloudiness and generate more thunderstorms and wildfires distally from the LIP. Latitudinal climate variability will enhance or impede the erosion process of continental LIPs and, therefore, the drawdown of atmospheric carbon dioxide and release of volatiles (van de Schootbrugge et al. 2020). Herein, we focus on the numerous and inter-related agents of catastrophic environmental change associated with LIPs and their impact on land plants, especially their manifestation in the palynological record as spore spikes, aberrant pollen and spores, reworked palynomorphs, and compositional changes that reflect large-scale reorganization of land plant communities.
 Spore Spikes as Evidence of Widespread Habitat Destruction
Spore Spikes as Evidence of Widespread Habitat Destruction
Crises at the end-Permian (Hochuli et al. 2010), end-Triassic (van de Schootbrugge et al. 2009; Lindström 2016), and end-Cretaceous (Vajda and Bercovici 2014), as well as smaller scale disturbances associated with LIP activity (Galloway et al. 2022), are all associated with increased abundances of spores of land plants (so-called “spore spikes”). Spore spikes, when referring to spores from land plants, reflect a floral response to major environmental disturbances, whereby opportunistic taxa, typically ferns, but also bryophytes (non-vascular plants such as mosses), lycophytes (vascular plants), and later in the geological record, angiosperms (flowering plants), colonize newly disturbed habitats. Fungal or algal spikes also occur, particularly at the end-Permian event, but are not further discussed herein (Dal Corso et al. 2022 and references therein). The spore spikes reflect pioneering plant communities, which, in the absence of renewed disturbance, are eventually replaced by successive intermediate species until a stable (“climax”) community is reached. This pattern of advancing ecological succession will be the same regardless of the cause of the disturbance, and spore spikes are often recorded in sedimentary successions, reflecting, for example, the first colonization of newly emerged areas during regressions. However, the spore spikes registered during mass extinction events are of a supra-regional to global extent and reflect substantial disturbance of the terrestrial ecosystem on larger scales (van de Schootbrugge et al. 2009). Several causal mechanisms often interact to increase the severity of the disruption, as was the case for the end-Triassic mass extinction where sea-level change, wildfire, and soil erosion led to habitat fragmentation and deforestation (van de Schootbrugge et al. 2009, 2020; Belcher et al. 2010; Lindström 2021).
The magnitude of the spore spike appears to be related to the severity of the crisis. Spore abundances during the end-Permian reached 95% in some locations (Hochuli et al. 2016). During the end-Triassic event, maximum spore abundances during the extinction interval reached 55%–90% of the spore–pollen floras (van de Schootbrugge et al. 2009; Lindström 2016). Fern, lycophyte, and bryophyte spores made up to 70% of the palynoflora in strata deposited after the first lava flows of the HALIP in Arctic Canada (Galloway et al. 2022). Fern pioneers are also well documented in response to smaller-scale disturbances associated with modern magmatic activity. For example, ferns were early colonizers of denuded ground after landslides associated with the 1980 Mount St. Helens eruption and were dominant plants following the cataclysmic eruption of Krakatau in 1883 (Tschudy et al. 1984). The dominance of ferns in particular following disturbance is the result of their tolerance of ecological stress, including their ability to grow on strongly leached and/or nutrient-poor or metal-enriched soils, tolerance of low-light conditions, short generation cycles, and other life-cycle traits such as gametophytic (haploid generation) self-fertilization and reproduction through segmentation of underground rhizomes (creeping rootstalks).
Wildfires
Wildfire is an integral component of land plant ecology. Low-intensity fires that burn close to the ground “clean” and “thin” the forest by removing thick flammable biomass from the forest floor, creating habitat for regeneration. In contrast, high-severity fires burn with intense heat and climb into and remove tree canopy, and combust soil organic matter and damage roots on the ground. Largescale magmatism can cause widespread wildfire (van de Schootbrugge et al. 2009; Belcher et al. 2010) via emplacement-related ignition or due to the higher atmospheric CO2 conditions that increase lightning strikes. Records of charcoal, fusinite (a maceral in coal), fly-ash, and polycyclic aromatic compounds (PAHs) indicate increased and intensified wildfires associated with many LIP and extinction events (Table 1). Following ignition, the frequency and severity of fires are related to a number of effects, including bottom-up controls such as vegetation, fuel, and topography, and top-down controls such as climate and the amount of atmospheric O2 (Belcher et al. 2010). Changes in fuel properties due to climate-induced vegetation shifts further modulate fire regimes (Belcher et al. 2010). Wildfire associated with LIPs affected vegetation in numerous and complex ways. Wildfires increased across the Paleocene– Eocene Thermal Maximum (PETM), even in high northern latitudes and even though conditions were humid, due to a climate-driven shift towards angiosperm-dominated vegetation that resulted in increased biomass and/or continuity of more fire-prone taxa (Denis et al. 2017). Across the end-Permian event, a spike in wildfire activity is implicated as one of many cumulative effects that caused abrupt extirpation of the primary coal-forming carbon sinks, such as the Glossopteris biome of Gondwana (Vajda et al. 2020) and the tropical gigantopterid and conifer forests of eastern Asia (Chu et al. 2020). Wildfires that occurred with the emplacement of CAMP during the end-Triassic event exacerbated widespread deforestation (van de Schootbrugge et al. 2009; Belcher et al. 2010) and contributed to catastrophic soil loss over extensive areas.
Reworked Palynomorphs as Indication of Large-scale Erosion
As a consequence of weathering and erosion processes, spores and pollen (and other palynomorphs) from older sedimentary units can be redeposited into younger sediments and mixed with the in-situ palynomorphs. During the end-Triassic event, catastrophic soil loss occurred across much of Europe as a consequence of deforestation from wildfires and acid rain, as well as seismic activity. This is evidenced by abundant reworked palynomorphs in sediments deposited during the crisis, both on land and at sea (van de Schootbrugge et al. 2020). A similar scenario has been proposed for the PETM (Korasidis et al. 2022). Such enhanced soil loss may have contributed to adverse conditions on the ocean shelves, causing eutrophication of surface waters through enhanced input of phosphorus and other bio-limiting nutrients that would have been delivered via massive-scale soil erosion and/ or transgression. Thus, what happens on land affects the marine environment as well (van de Schootbrugge et al. 2020).
Sea-level Change
Sea-level changes caused by LIP emplacement have also influenced land plants over geological time. At the end-Triassic event, associated with emplacement of the CAMP, climate and sea-level changes caused fragmentation and destruction of coastal and near-coastal lowland mire habitats around the European epicontinental sea and Tethys margin (Lindström 2021). This habitat change, together with other ecological stressors induced by volcanism, caused a reduction in abundance and geographic range of gymnosperm trees and shrubs adapted to those environments. A subsequent sea-level fall then promoted colonization of disturbed areas by pioneering opportunists and herbaceous survivors in an environment already stressed by increased wildfire and enhanced soil erosion. Sea-level rise ultimately restored some of the near-coastal mire habitats and recovery ensued. In another illustration of complex feedbacks, water table rise, as a response to the abrupt disappearance of vegetation from the landscape, during and following the end-Permian event, increased salinity that further compromised terrestrial habitability (Vajda et al. 2020).
Spore-pollen Mutagenesis—Evidence of Ozone Depletion, Hg-pollution, or Both?
Volcanic activity can emit a range of known primary phytotoxic pollutants, including Hg and other heavy metals, fluoride, O3, SO2, and PAHs. For instance, lesions on fossil plant leaves indicate that plants were subjected to SO2-induced damage from acid rain during the end-Triassic crisis (Steinthorsdottir et al. 2018). Phytotoxic substances can induce stress responses and cause morphologically visible changes (e.g., stunted growth, lesions, necrosis, root shortening) in the parent plants, but can also cause damage leading to mutations in spores and pollen. While various common environmental stresses for plants (e.g., drought, frost, water logging) can disrupt the spore or pollen maturation processes and result in premature shedding of immature spores or pollen that may or may not be retained and dispersed in tetrads, these disruptions are generally local and short-lived from a geological point of view. One of the most striking features in the palynological record contemporaneous with several LIP-associated extinction events is the presence of mutated spores and pollen (Fig. 1; Table 1). Based on both living and fossil plants, normal spore/pollen formation results in 95%–97% viable and 3%–5% underdeveloped or abnormal, non-viable spores or pollen. Aberrant spore/pollen quantities above 5% are, therefore, indicative of environmental stress (Lindström et al. 2019). Lindström et al. (2019) document an increase of up to 70% aberrant spores at the end-Triassic extinction event in the Danish and German basins contemporaneous with an increase in Hg and a negative carbon isotope excursion (see Title Figure). During the Smithian–Spathian boundary event (a late Early Triassic crisis in the wake of the end-Permian event), ~50% of spores and pollen were “malformed” across the negative δ13Corg shift of this event in Pakistan (Galasso et al. 2022); and during the end-Permian crisis, 40% of the total assemblage co-occurring with the onset of the negative δ13Corg shift were considered abnormal, indicating that the reproductive ability of the parent plants was inhibited (Hochuli et al. 2017). Spore/pollen mutagenesis during LIP events could have been caused by increased and lethal UV-B radiation as a result of stratospheric ozone depletion by halocarbon and aerosol emissions from LIP magmatism (Benca et al. 2018, 2022), volcanic induced Hg-toxicity (or other heavy metals) (Lindström et al. 2019), and possibly acid rain (SO2) and soil acidification (Hochuli et al. 2017; van de Schootbrugge et al. 2009) in addition to general environmental stress.
Not all LIP emissions were deleterious to land plants. Mercury is toxic to many plant species but not to all: it can damage cell division, is mutagenic, but can also induce polyploidization that is adaptive in some cases. Loading of atmospheric gaseous Hg associated with LIP activity could have contributed to ecological turnover of terrestrial vegetation by damaging the development of some species while benefitting others (Lindström et al. 2019). Phosphorus is emitted from volcanism and thermal fixation (reaction of P with other minerals to form insoluble compounds, facilitated by heat) and subsequent atmospheric oxidation of NOx in volcanic environments provide essential macronutrients to land plants (Jolley et al. 2008). Phosphorus levels are further enhanced from weathering and breakdown of soil minerals, basaltic lava, and ash. Nitrogen becomes available by decomposition and burning of already-established biomass as well as NO3 production at the interface between hot lava (thermal fixation) and the atmosphere (Mather et al. 2004). Eutrophication from phosphate and nitrate deposition derived from volcanic gases and weathering of volcanic products may have provided conditions that favored development of high-productivity ecosystems during LIP emplacement (Jolley et al. 2008). Alkali-rich volcanic ash from LIPs could also have supplied nutrients (e.g., K, Ca, Na) to land plants.
Climate Change
Warming is linked to synergistic stresses for land plants and is likely part of the “lethal cocktail” of LIP effects (Bond and Sun 2021). After the end-Permian mass extinction, temperatures on land may at times have reached >40 °C during the late Smithian Thermal Maximum (Sun et al. 2012), which is too hot for most land plants to survive. This undoubtedly contributed to the prolonged recovery of terrestrial ecosystems at this time. The Jenkyns Event was another hyperthermal episode of the Phanerozoic, associated with a second-order extinction event in the early Toarcian (ca. 183 Ma; Early Jurassic) and associated, in part, with outgassing from emplacement of the Karoo-Ferrar LIP. While the Jenkyns Event is manifested mostly as changes in marine ecosystems, it is also recorded in terrestrial deposits by changes in the diversity and composition of land plants. Slater et al. (2019) examined pollen and spore assemblages from Tethyan Pliensbachian–Toarcian strata and showed that vegetation across the Jenkyns Event shifted from high diversity mixed conifers, seed ferns, and wet-adapted ferns and lycophytes to an assemblage dominated by plants adapted to drier conditions, such as cheirolepids and other conifers. Thus, the event caused a substantial short-term reduction in floral richness and diversity, but also a longerterm turnover in terrestrial vegetation. In Arctic Canada, an interval of Hauterivian to early Barremian warming related to recovery from the Valanginian cold snap, and/or CO2 forced warming associated with Paraná-Etendeka Province activity, resulted in an expansion of hinterland communities dominated by Pinaceae (Galloway et al. 2022). The PETM was a hyperthermal event associated with emplacement of the NAIP that caused changes in both marine and terrestrial realms through effects on seasonality and precipitation (Jaramillo et al. 2010; Korasidis et al. 2022).
Magmatic activity of LIPs increased the frequency and intensity of climate extremes, such as prolonged heatwaves, drought, precipitation changes, fluctuating groundwater, major storms, and wildfires; and also caused atmospheric pollution through volcanic dust, sulfur dioxide, and heavy metals, which contributed to major biotic crises in the past. Hence, the geological record of LIP-induced extinction events provides an informative archive on how land plants may respond to future environmental stress.
CONCLUSIONS
Large-scale magmatic events set off a biogeochemical cascade that affects the atmosphere, geosphere, biosphere, and hydrosphere of the planet. LIPs occur contemporaneous with almost all major biotic crises throughout the history of life on Earth. While devastation to animal life appears to have been greater than effects on land plants, LIPs nonetheless have provoked major changes in terrestrial vegetation. Specific and interacting agents of vegetation change include habitat destruction, wildfire, climate change, sea-level change, and emissions of toxic substances that are variously manifested in the pollen and spore record. Spore spikes reflect habitat disturbance, compositional changes, abnormal pollen and spores as a result of environmental stress and mutagenesis, and abundant reworked pollen and spores that reflect soil erosion on a massive scale. These causal agents interact with each other; for example, climate change induces vegetation changes that, in turn, affect wildfire intensity and frequency. Understanding the response of land plants to major biotic crises, and the complex feedbacks associated with vegetation change, can inform adaptive mitigation strategies in the face of current and forecasted climate change.
ACKNOWLEDGMENTS
This publication represents NRCan Contribution Number/ Numéro de contribution de RNCan 20220527. JMG received funding from the AIAS-COFUND II fellowship programme that is supported by the Marie Skłodowska-Curie actions under the European Union’s Horizon 2020 (grant agreement no. 754513) and the Aarhus University Research Foundation for part of this work. We thank Dr. Stephen Grasby for an internal peer review. We thank Daoliang Chu and Jun Shen as external peer reviewers and the Editorial Team whose comments greatly improved our manuscript.
REFERENCES
Belcher CM and 7 coauthors (2010) Increased fire activity at the Triassic/Jurassic boundary in Greenland due to climatedriven floral change. Nature Geoscience 3: 426-429, doi: 10.1038/ngeo871
Benca JP, Duijnstee, IAP, Looy CV (2018) UV-B–induced forest sterility: implications of ozone shield failure in Earth’s largest extinction. Science Advances 4: e1700618, doi: 10.1126/sciadv.1700618
Benca JP, Duijnstee IA, Looy CV (2022) Fossilized pollen malformations as indicators of past environmental stress and meiotic disruption: insights from modern conifers. Paleobiology 48, 677-710, doi: 10.1017/pab.2022.3
Bond DPG, Sun Y (2021) Global warming and mass extinctions associated with large igneous province volcanism. In: Ernst RE, Dickson AJ, Bekker A (eds) Large Igneous Provinces: A Driver of Global Environmental and Biotic Changes, Volume 255 (First Edition). American Geophysical Union, John Wiley and Sons Inc, Washington DC, pp 83-102, doi: 10.1002/9781119507444.ch3
Chu D and 10 coauthors (2020) Ecological disturbance in tropical peatlands prior to marine Permian–Triassic mass extinction. Geology 48: 288-292, doi: 10.1130/ G46631.1
Dal Corso J and 8 coauthors (2022). Environmental crises at the Permian– Triassic mass extinction. Nature Reviews Earth & Environment 3: 197-214, doi: 10.1038/s43017-021-00259-4
Denis EH, Pedentchouk N, Schouten S, Pagani M, Freeman KH (2017) Fire and ecosystem change in the Arctic across the Paleocene–Eocene Thermal Maximum. Earth and Planetary Science Letters 467: 149-156, doi: 10.1016/j.epsl.2017.03.021
Galasso F, Bucher H, Schneebeli-Hermann E (2022) Mapping monstrosity: malformed sporomorphs across the Smithian/Spathian boundary interval and beyond (Salt Range, Pakistan). Global and Planetary Change 219: 103975, doi: 10.1016/j. gloplacha.2022.103975
Galloway JM and 7 coauthors (2022) Exploring the role of High Arctic Large Igneous Province volcanism on Early Cretaceous Arctic forests. Cretaceous Research 129: 105022, doi: 10.1016/j. cretres.2021.105022
Hochuli PA, Hermann E, Vigran JO, Bucher H, Weissert H (2010) Rapid demise and recovery of plant ecosystems across the end-Permian extinction event. Global and Planetary Change 74: 144-155, doi: 10.1016/j.gloplacha.2010.10.004
Hochuli, PA, Sanson-Barrera, A, SchneebeliHermann E, Bucher H (2016) Severest crises overlooked—worst disruption of terrestrial environments postdates the Permian–Triassic mass extinction. Scientific Reports 6: 28372, doi: 10.1038/ srep28372
Hochuli PA, Schneebeli-Hermann E, Mangerud G, Bucher H (2017) Evidence for atmospheric pollution across the Permian– Triassic transition. Geology 45: 1123-1126, doi: 10.1130/G39496.1
Jaramillo C and 28 coauthors (2010). Effects of rapid global warming at the Paleocene– Eocene boundary on neotropical vegetation. Science 330: 957-961, doi: 10.1126/ science.1193833
Jolley DW, Widdowson W, Self S (2008) Volcanogenic nutrient fluxes and plant ecosystems in large igneous provinces: an example from the Columbia River Basalt Group. Journal of the Geological Society 165: 955-966, doi: 10.1144/0016-76492006-199
Korasidis VA, Wing SL, Shields CA, Kiehl JT (2022) Global changes in terrestrial vegetation and continental climate during the Paleocene–Eocene Thermal Maximum. Paleoceanography and Paleoclimatology 37: e2021PA004325, doi: 10.1029/2021PA004325
Lindström S (2016) Palynofloral patterns of terrestrial ecosystem change during the end-Triassic event–a review. Geological Magazine 153: 223-251, doi: 10.1017/ S0016756815000552
Lindström S (2021) Two-phased mass rarity and extinction in land plants during the end-Triassic climate crisis. Frontiers in Earth Science 9: 780343, doi: 10.3389/ feart.2021.780343
Lindström S and 8 coauthors (2019) Volcanic mercury and mutagenesis in land plants during the end-Triassic mass extinction. Science Advances 5: eaaw4018, doi: 10.1126/sciadv.aaw4018
Lindström S and 8 coauthors (2021) Tracing volcanic emissions from the Central Atlantic Magmatic Province in the sedimentary record. Earth-Science Reviews 212: 103444, doi: https://doi.org/10.1016/j. earscirev.2020.103444
Mather TA (2015) Volcanoes and the environment: lessons for understanding Earth’s past and future studies of present-day volcanic emissions. Journal of Volcanology and Geothermal Research 304: 160-179, doi: 10.1016/j.jvolgeores.2015.08.016
Mather TA, Pyle DM, Allen AG (2004) Volcanic source for fixed nitrogen in the early Earth’s atmosphere. Geology 32: 905-908, doi: 10.1130/G20679.1
Nowak H, Schneebeli-Hermann E, Kustatscher E (2019) No mass extinction for land plants at the Permian–Triassic transition. Nature Communications 10: 384, doi: 10.1038/s41467-018-07945-w
Slater SM, Twitchett RJ, Danise S, Vajda V (2019) Substantial vegetation response to Early Jurassic global warming with impacts on oceanic anoxia. Nature Geoscience 12: 462-467, doi: 10.1038/s41561-019-0349-z
Steinthorsdottir M, Elliott-Kingston C, Bacon KL (2018) Cuticle surfaces of fossil plants as a potential proxy for volcanic SO2 emissions: observations from the Triassic–Jurassic transition of East Greenland. Palaeobiodiversity and Palaeoenvironments 98: 49-69, doi: 10.1007/s12549-017-0297-9
Sun Y and 7 coauthors (2012) Lethally hot temperatures during the Early Triassic greenhouse. Science 338: 366-370, doi: 10.1126/science.1224126
Tschudy RH, Pillmore CL, Orth CJ, Gilmore JS, Knight JD (1984) Disruption of the terrestrial plant ecosystem at the Cretaceous–Tertiary boundary, Western Interior. Science 225: 1030-1032, doi: 10.1126/science.225.4666.1030
Vajda V, Bercovici A (2014) The global vegetation pattern across the Cretaceous– Paleogene mass extinction interval: a template for other extinction events. Global and Planetary Change 122: 29-49, doi: 10.1016/j.gloplacha.2014.07.014
Vajda V and 8 coauthors (2020) End-Permian (252 Mya) deforestation, wildfires and flooding—an ancient biotic crisis with lessons for the present. Earth and Planetary Science Letters 529: 115875, doi: 10.1016/j. epsl.2019.115875
van de Schootbrugge B and 11 coauthors (2009) Floral changes across the Triassic/ Jurassic boundary linked to flood basalt volcanism. Nature Geoscience 2: 589-594, doi: 10.1038/ngeo577
van de Schootbrugge B and 11 coauthors (2020) Catastrophic soil loss associated with end-Triassic deforestation. Earth-Science Reviews 210: 103332, doi: 10.1016/j.earscirev.2020.103332